|
This section contains 1,124 words (approx. 4 pages at 300 words per page) |

|
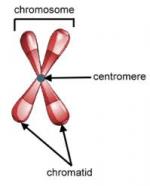
Chromosome segregation refers to the coordinated movement of chromosomes to opposite poles of the cell during either cellular reproduction (mitosis) or the production of sex cells (meiosis). Chromosomes segregate along the spindle apparatus, a microtubule-based structure that attaches at a site called the kinetochore to the chromosomes centromere during anaphase of mitosis. To ensure equal segregation, sister chromatids are linked together from the time of their synthesis (S-phase of the cell cycle) to the time of their segregation. Without segregation of chromosomes, there would be no way to pass on genetic information from one cell to the next. This process, however, is not flawless. Chromosome abnormalities can arise when chromosomes undergo rearrangements or fail to segregate properly. These alterations can be of a structural or numerical nature. Numerical anomalies result in aneuploidy, or addition or loss of individual chromosomes from the normal...
|
This section contains 1,124 words (approx. 4 pages at 300 words per page) |
|



