|
This section contains 1,450 words (approx. 5 pages at 300 words per page) |

|
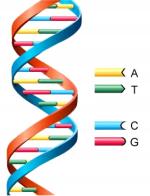
Deoxyribonucleic acid (DNA) is a double-stranded, helical molecule that forms the molecular basis for heredity.
For replication (duplication) to occur, DNA must first unwind, or "unzip," itself to allow the genetic information-encoding bases to become accessible. The base pairing within DNA is specific and complementary and, consequently, when the molecule unwinds, two complimentary strands are temporarily produced, each of which acts as a template for a new strand. At the onset of replication, a replication fork is created as the DNA molecule separates at a small region. The enzyme DNA polymerase then adds complementary nucleotides to each side of the freshly separated strands. The DNA polymerase adds nucleotides only to one end of the DNA. As a result, one strand (the leading strand) is replicated continuously, while the other strand (the lagging strand) is replicated discontinuously, in short bursts. Each of these small sections...
|
This section contains 1,450 words (approx. 5 pages at 300 words per page) |
|



